
Опишите процесс заселения песчаной дюны биология
6.4. Экологическая сукцессия. Классификация сукцессии. Проблемы стабильности экосистем
Напомним, что под экологической сукцессией понимается постепенная необратимая направленная смена одних биоценозов другими на одной и той же территории под влиянием природных факторов или воздействия человека.
Наблюдать сукцессию можно на заброшенных полях разного возраста, песчаных дюнах или песчаных морских и речных берегах. Если рассматривать сукцессию на брошенных землях, которые не используются в сельском хозяйстве, то можно заметить, что бывшие поля быстро покрываются разнообразными однолетними растениями (рис. 6.14). Сюда же попадают семена древесных пород: сосны, ели, березы, осины. Они легко и на большие расстояния разносятся ветром, а также животными. Попав на слабозадерненную почву, семена начинают прорастать, причем в наиболее благоприятном положении оказываются мелколиственные породы (береза, осина). Хвойные обычно гибнут из-за бурного развития трав, под влиянием разлагающегося опада или от обилия минеральных элементов.
Вначале изменения происходят быстро, затем скорость сукцессии снижается по мере появления растений, развивающихся

Рис. 6.14. Сукцессия: формирование елового леса на брошенных землях

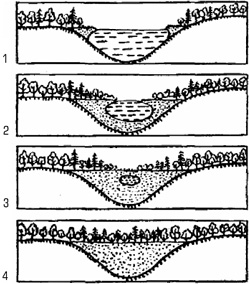
Рис. 6.15. Превращение озера в низинное болото
на болотах начинается с того, что края водоема зарастают водными растениями. Вблизи берегов сплошным ковром разрастаются влаголюбивые виды растений (камыш, тростник, осока). Создается более или менее плотный слой растительности на поверхности воды. Отмершие остатки растений постепенно накапливаются на дне водоема. Из-за малого количества кислорода в застойных водах эти остатки медленно разлагаются и превращаются в торф. Начинается формирование болотного биоценоза: появляются сфагновый мох, на сплошном ковре которого поселяются клюква, багульник, голубика, кое-где сосенки, образующие редкую поросль тощих деревьев. Так с течением времени постепенно образуется биоценоз верхового болота.
Все сукцессии можно разделить на две большие группы. Это прежде всего аллогенные сукцессии (от греч. alios - иной, другой и genesis - возникновение), причины которых определяются внешними влияниями (природными или антропогенными), изменяющими условия среды. Таким образом, при ал- логенной сукцессии источник изменения биоценоза находится в окружающей его среде. Движущие силы сукцессии имеют направленный характер: среда → растительность. К такому типу сукцессий можно отнести переход от эвтрофного озера, обогащенного питательными веществами стоков (например, животноводческой фермы), к болоту или наземному сообществу.
Сукцессии, происходящие в результате изменения условий среды самими сообществами в отсутствие постепенного

Рис. 6.16. Последствия взрыва вулкана Кракатау:
1 - зона распространения звуковой волны от взрыва; 2 - площадь пеллопада; 3 - вулкан Кракатау
изменения абиотических факторов, называются автогенными (от греч. autos - сам и genesis - происхождение). В данном случае источником смены служит сама растительность, которая посредством изменения среды своего существования меняет структуру. Происходит как бы «самоотрицание» растительности. Схематично этот процесс можно представить следующим образом: растительность → среда → растительность. Автогенные сукцессии могут быть первичными: развитие сообществ идет во вновь образовавшихся местообитаниях, на новых субстратах, где растительность ранее отсутствовала, - на песчаных дюнах, застывших потоках лавы, на породах, обнажившихся в результате эрозии или отступления льдов.
Интересным примером первичной сукцессии может служить заселение живыми существами острова Кракатау, расположенного между островами Суматра и Ява в Индонезии. Ранним августовским утром 1883 г. взорвалась вершина вулкана Кракатау. Его взрывное извержение и последовавшая за этим мощная волна цунами высотой до 40 м унесли жизни свыше 36 тыс. человек. Звук взрыва тогда был слышен на расстоянии 4650 км, грохот разорвал барабанные перепонки моряков за 40 км от него, а атмосферные эффекты наблюдались по всему миру (рис. 6.16).
На воздух было поднято 24 км3 скал. Горячий пепел засыпал территорию площадью свыше 800 тыс. км2. От острова осталась только гора, покрытая пеплом. Интенсивные зори, возникшие в результате уменьшения прозрачности

Рис. 6.17. Возрождение жизни на Кракатау
атмосферы, наблюдались здесь в течение нескольких лет. Все живое было погребено под потоками горящей лавы. Не осталось никаких признаков жизни. Даже мельчайшие споры и семена растений оказались в плену разбушевашейся стихии. Перед учеными открылась перспектива изучения развития жизни при естественном заселении субстрата голых скал, возникшего после извержения.
Процесс заселения на острове шел несколькими путями (рис. 6.17).
Большая часть растений и животных прибыла с Явы и Суматры, расположенных примерно в 40 км от Кракатау. Первым на острове через 9 месяцев после взрыва был обнаружен плетущий паутину паук (1). Занесенные ветрами синезеленые водоросли подготовили условия, в которых уже могли прорастать микроскопические споры мхов, папоротников (2) и таких цветковых растений, как эмилия (3) и веделня (4). Море выбросило на берег семена ка- лофиллума (5), турнефорции (б) и ипомеи (7): все они проросли и пустили корни. По мере того как формировался почвенный покров, растениям становилось все легче обживаться на острове. В 1896 г. занесенные ветром семена дикого сахарного тростника дали буйные побеги, и даже крохотные спороподобные семена прихотливой орхидеи (5) нашли себе подходящее пристанище.

Рис. 6.18. Динамика видового разнообразия на острове Кракатау после полного уничтожения аборигенной флоры и фауны вулканическим взрывом в 1893 г.:
крестики - число видов растений; светлые кружки - число видов птиц; черные кружки - прочие виды
Проросли и заброшенные прибоем выше верхней отметки прилива семена таких тропических деревьев, как барринггония (9), казуарина (10) и кокосовая пальма (11). Случайные птицы и насекомые, вероятно, довольно часто попадали на остров и, по мере того как появлялись необходимые для их существования растения, оставались здесь. В пищеварительном тракте птиц на остров заносились семена фигового дерева (12) и других растений. Некоторые семена попадали сюда, прицепившись к перьям птиц. Если не считать таких отличных пловцов, как питон (13) и варан (14), большинство пресмыкающихся - гекконы (15), агамы (16) и сцинки или их яйца - попадали на остров в основном на плавающих деревьях.
По прошествии четверти века на острове насчитывалось уже 263 вида животных. Помимо обилия насекомых, здесь поселились 16 видов птиц, 2 вида пресмыкающихся и 4 вида улиток. Позже были занесены гекконы, агамы, а отличные пловцы питоны и вараны добрались своим ходом. Спустя 50 лет на Кракатау появился молодой, правда, низкорослый, лес, а фауна уже насчитывала около 1200 видов (рис. 6.18). Таким образом, заселение живыми существами острова Кракатау является примером первичной сукцессии.
Если на какой-либо местности ранее существовала растительность, но по каким-либо причинам она была уничтожена, то ее естественное восстановление называется вторичной сукцессией. К таким сукцессиям может привести, например, локальное уничтожение леса болезнями, ураганом, извержением вулкана, землетрясением - так называемая катастрофическая сукцессия, либо пожаром - пирогенная сукцессия. Большинство сукцессий, наблюдаемых в настоящее время, являются сукцессиями антропогенными (от греч. anthropos - человек). Это выпас скота, рубка лесов, возникновение очагов возгорания, распашка земель, затопление почв, опустынивание и т.п.
Каждая стадия сукцессии характеризуется изменениями не только в структуре растительности, но и в животном компоненте биоценоза. По мере роста древостоя животное население в значительной степени меняет свой состав. Появляющиеся хищники и паразиты контролируют видовую структуру биоценоза. Поэтому последовательная и непрерывная смена видов во времени является характерной чертой большинства сукцессионных процессов. Сукцессии характерны как для сухопутных, так и для водных местообитаний. Их можно наблюдать и в мелководных экосистемах (озеро, пруд), и в крупных реках, и даже в океане. Обычно на процессы сукцессии в водных местообитаниях указывают изменения в составе прибрежной растительности и обилии водных животных, главным образом рыбы. Часто при строительстве водохранилищ приходится затапливать водой большие площади суши с более или менее плодородными почвами. Обилие биогенных элементов на этой стадии приводит к вспышке численности рыбы, количество которой затем снижается по мере расходования запасов пищи. В дальнейшем происходит стабилизация обилия рыбного населения, поддерживающаяся определенным уровнем содержания питательных веществ.
Рассматривая сукцессии растительности при создании водохранилищ, можно установить некоторые общие закономерности. Так, во всех случаях формируются сообщества свободноплавающих, прикрепленных растений и прибрежно-водные группировки из рогозов, манника, ежеголовника, тростника и т.д. Такая стадийность зарастания водных местообитаний характерна для многих водных экосистем, однако некоторые стадии могут выпадать в зависимости от конкретных природных условий.
По мере развития биоценоза, происходящего в результате смены различных сукцессионных стадий, изменения его структуры и видового состава протекают до определенного предела, после которого сообщество приходит в относительное стабильное состояние, главным образом за счет стабилизации структуры растительности. Такое относительно устойчивое и равновесное по отношению к внешней среде растительное сообщество носит название климаксового. Таким образом, климакс (от греч. klimax - лестница) представляет собой заключительную стадию развития биоценоза, на которой он находится в равновесном состоянии с окружающей средой довольно продолжительное время.
В таких климаксовых сообществах долгое время сохраняются относительно постоянные потоки вещества и энергии. Видовой состав этих сообществ обычно богат и однороден по структуре. В течение сукцессии биомасса и продукция живых организмов возрастает, а круговорот веществ становится все более замкнутым.
Стабильность экосистемы возможна только при сбалансированном поступлении и отчуждении органического вещества. В этом случае общая живая масса экосистемы остается в устойчивом состоянии. Под экологической стабильностью экосистемы понимается способность ее противостоять внутренним и внешним абиотическим и биотическим факторам среды.
Глава 2. Сукцессия
Происходящие изменения в сообществах экосистемы приводят, в конечном счете, к замене одного сообщества другим, с иным набором видов — доминантов. Причиной подобных замен могут быть внешние факторы, длительное время действующие в одном направлении: мелиоративное осушение болотных почв, загрязнение водоемов, перевыпас пастбищ и др. Если при этом усиливающееся влияние фактора приводит к постепенному упрощению структуры сообществ, обеднению их состава, снижению продуктивности, то подобные смены называют дигрессионными.Закономерный направленный процесс изменения сообществ в результате взаимодействия живых организмов между собой и окружающей их абиотической средой называют сукцессией, то есть, направленная смена одних биоценозов другими на одной и той же территории под влиянием природных факторов или воздействия человека.
Экологические сукцессии - закономерное направленное развитие экосистемы, при котором происходит смена во времени одного сообщества другим. Смена сообществ базируется на функциях автотрофов. Гетеротрофная составляющая экосистемы формируется на базе фитоценозов и лишь вторично начинает влиять на его состав и свойства. Смена последовательного ряда сообществ – адаптивный ответ на экосистемном уровне по отношению к изменяющимся факторам среды (климат, рельеф, гидролог. режим).
Сукцессия завершается образованием стабильного сообщества, наиболее адаптированного к данным сложившимся условиям – климакс. В определенных условиях сукцессия может быть регрессивной – обеднение и упрощение сообществ, например, в результате антроп. воздействий.
Наблюдать сукцессию можно на заброшенных полях раннего возраста, песчаных дюнах или песчаных морских и речных берегах. Если мы будем рассматривать сукцессию на брошенных землях, которые не используются в сельском хозяйстве, то можно видеть, что бывшие поля быстро покрываются разнообразными однолетними растениями. Сюда же могут попасть, преодолев иногда большие расстояния с помощью ветра или животных, семена древесных пород: сосны, ели, березы, осины.
Вначале изменения происходят быстро. Затем, по мере появления растений, растущих более медленно, скорость сукцессии снижается. Всходы березы образуют густую поросль, которая затеняет почву, и даже если вместе с березой прорастают семена ели, ее всходы, оказавшись в весьма неблагоприятных условиях, сильно отстают от березовых. Светолюбивая береза является серьезным конкурентом для ели. К тому же специфические биологические особенности березы дают ей преимущества в росте. Березу называют «пионером леса», так как она почти всегда первой поселяется на нарушенных землях и обладает широким диапазоном приспособляемости.
Березки в возрасте 2—3 лет могут достигать высоты 100—120 см, тогда как елочки в том же возрасте едва дотягивают до 10 см. Постепенно, к 8—10 годам березы формируют устойчивое березовое насаждение высотой до 10—12 м. Под развивающимся пологом березы начинает подрастать и ель, образуя разной степени густоты подрост. Перемены происходят и в нижнем, травяно-кустарничковом ярусе. Постепено, по мере смыкания крон березы, светолюбивые виды, характерные для начальных стадий сукцессии, начинают исчезать и уступают место теневыносливым.

Изменения касаются и животного компонента рассматриваемого биоценоза. На первых стадиях поселяются майские хрущи, березовые пяденицы, затем появляются многочисленные птицы: зяблики, славки, пеночки. Поселяются мелкие млекопитающие: землеройки, кроты, ежи. Изменение условий освещения начинает благоприятно сказываться на молодых елочках, которые ускоряют свой рост. Если на ранних этапах сукцессии прирост елочек составил 1—3 см в год, то по прошествии 10—15 лет он достигает уже 40—60 см. Где-то к 50 годам ель догоняет березу в росте, и образуется смешанный елово-березовый древостой. Из животных появляются зайцы, лесные полевки и мыши, белки. Заметны сукцессионные процессы и среди птичьего населения. Появляются иволги, питающиеся гусеницами.
Смешанный елово-березовый лес постепенно сменяется лесом еловым. Ель перегоняет в росте соперницу-березу, создает значительную тень, и светолюбивая белоствольная красавица, не выдержав конкуренции, постепенно выпадает из древостоя. Таким образом происходит сукцессия, при которой вначале березовый, затем смешанный елово-березовый лес сменяется чистым ельником. Естественный процесс смены березняка ельником длится более 100 лет. Именно поэтому иногда процесс сукцессии называют вековой сменой.

Если развитие сообществ идет на вновь образовавшихся, ранее никем и ничем не заселенных местообитаниях, — на песчаных дюнах, застывших потоках лавы, породах, обнажившихся в результате эрозии или отступления льдов, то такая сукцессия называется первичной.
Так, в первичных сукцессиях, протекающих на скалах Уральских гор, различают следующие этапы.
1. Поселение эндолитических и накипных лишайников, сплошь покрывающих каменистую поверхность. Накипные лишайники несут своеобразную микрофлору и содержат богатую фауну простейших, коловраток, нематод. Мелкие клещи — сапрофаги и пер-вичнобескрылые насекомые обнаруживаются сначала только в трещинах. Активность всего населения прерывиста, отмечается главным образом после выпадения осадков в виде дождя или смачивания скал влагой туманов. Данные сообщества организмов называют пионерными.
2. Преобладание листоватых лишайников, которые постепенно образуют сплошной ковер. Под круговинками лишайников в результате выделяемых ими кислот и механического сокращения слоевищ при высыхании образуются выщербленности, идет отмирание слоевищ и накопление детрита. В большом количестве под лишайниками встречаются мелкие членистоногие: коллемболы, панцирные клещи, личинки комаров-толкунчиков, сеноеды и другие. Образуется микрогоризонт, состоящий из их экскрементов.
3. Поселение литофильных мхов Hedwidia u Pleurozium schreberi. Под ними погребаются лишайники и подлишайниковые пленочные почвы. Ризоиды мхов здесь прикрепляются не к камню, а к мелкозему, который имеет мощность не менее 3 см. Колебания температуры и влажности под мхами в несколько раз меньше, чем под лишайниками. Усиливается деятельность микроорганизмов, увеличивается разнообразие групп животных.
4. Появление гипновых мхов и сосудистых растений. В разложении растительных остатков и формировании почвенного профиля постепенно уменьшается роль мелких членистоногих и растет участие более крупных беспозвоночных — сапрофагов: энхитреид, дождевых червей, личинок насекомых.
5. Заселение крупными растениями, способствующее дальнейшему накоплению и образованию почвы. Ее слой оказывается достаточным для развития кустарников и деревьев. Их опадающие листья и ветви не дают расти мхам и большинству других мелких видов, начавших сукцессию. Так, постепенно на изначально голых скалах идет процесс смены лишайников мхами, мхов травами и наконец лесом. Такие сукцессии в геоботанике называют экогенетически-ми, так как они ведут к преобразованию самого местообитания.
В качестве примера первичной сукцессии приведем процесс заселения вновь образованных песчаных дюн, где растительность прежде отсутствовала. Здесь вначале поселяются многолетние растения, способные переносить засушливые условия. Они укрепляют поверхность дюны и обогащают песок органическими веществами. Вслед за многолетниками появляются однолетники. Их рост и развитие часто способствуют обогащению субстрата органическим материалом, так что постепенно создаются условия, подходящие для произрастания таких растений, как ива, толокнянка, чабрец. Эти растения предшествуют появлению проростков сосны, которые закрепляются здесь и, подрастая, образуют через много поколений сосновые леса на песчаных дюнах.
Примером первичной сукцессии является также возрождение жизни на залитых потоками лавы склонах вулкана.
Примером вторичной сукцессии является рассмотренное выше зарастание заброшенного поля. К таким сукцессиям может привести, например, частичное уничтожение леса болезнями, ураганом, землетрясением либо пожаром. Большинство сукцессии, наблюдаемых в настоящее время, являются сукцессиями антропогенными (от греч. anthropos — человек), т.е. происходящими в результате воздействия человека на природные экосистемы. Это выпас скота, рубка лесов, возникновение очагов возгорания, распашка земель, затопление почв, опустынивание и т.п.
Примером вторичной сукцессии может служить образование торфяного болота при зарастании озера (рис. 2.12). Изменение растительности на болотах начинается с того, что края водоема зарастают водными растениями. Влаголюбивые виды (камыштростник, осока) начинают разрастаться вблизи берегов сплошным ковром. Постепенно создается более или менее плотный слой растительности на поверхности воды. Отмершие остатки растений постепенно накапливаются на дне водоема. Из-за малого количества кислорода в застойных водах погибшие растения медленно разлагаются и постепенно превращаются в торф. Начинается формирование болотного биоценоза. Появляются сфагновые мхи, на сплошном ковре которых поселяются клюква, багульник, голубика. Здесь же могут поселяться сосенки, образуя редкую поросль тощих деревьев. Постепенно, с течением времени, образуется экосистема верхового болота.
Ни один вид растений или животных не может процветать на протяжении всей сукцессии. По мере роста древостоя состав животного населения во многом меняется. Появляющиеся хищники и паразиты в значительной степени контролируют видовую структуру биоценоза. Поэтому последовательная и непрерывная смена видов во времени является характерной чертой большинства сукцессионных процессов.
При развитии биоценоза изменения его структуры и состава видов протекают до определенного предела, после которого наступает относительно стабильное состояние сообщества, главным образом за счет стабилизации структуры растительности. Такое относительно устойчивое и равновесное по отношению к внешней среде растительное сообщество носит название климаксового сообщества. Таким образом,климакс (от греч. klimax — лестница) представляет собой заключительную стадию развития биоценоза, на которой он находится в равновесном состоянии с окружающей средой довольно продолжительное время.
В течение сукцессии биомасса живых организмов возрастает, а круговорот веществ увеличивается.
СОДЕРЖАНИЕ
Экология СПРАВОЧНИК
На кривой наклономера в песчаной дюне (рис.4-32), каждая граница передовых тонких слоев более или менее наклонена, как можно видеть на обнажении. Можно также определить направление ветра для каждого отложения. См. также изображение Formation MicroScanner на рис.4-33.[ . ]
| Возрастной состав клонов песчаной осоки Carex arenaria, растущих на песчаных дюнах Северного Уэльса (клоны состоят из разновозрастных побегов). Внесение минеральных удобрений приводит к тому, что этот возрастной состав изменяется |  |
Описательные исследования сукцессии песчаных дюн, степей, лесов, морских побережий или других участков наряду с более новыми функциональными исследованиями позволили построить теорию, лежащую в основе приведенных выше определений. Г. Одум и Пинкертон (1955), опираясь на сформулированное Лоткой (1925) «правило максимума энергии в биологических системах», впервые указали на то, что сукцессия связана с фундаментальным сдвигом потока энергии в сторону увеличения количества энергии, направленного на поддержание системы. Позднее Маргалеф (1963, 1968) подкрепил эту концепцию о биоэнергетических основах сукцессии фактическими данными.и расширил ее.[ . ]
Описательные исследования сукцессии песчаных дюн, степей, лесов, морских побережий или других участков наряду с более новыми функциональными исследованиями позволили построить теорию, лежащую в основе приведенных выше определений. Г. Одум и Пинкертон (1955), опираясь на сформулированное Лоткой (1925) «правило максимума энергии в биологических системах», впервые указали на то, что сукцессия связана с фундаментальным сдвигом потока энергии в сторону увеличения количества энергии, направленного на поддержание системы. Позднее Маргалеф (1963, 1968) подкрепил эту концепцию о биоэнергетических основах сукцессии фактическими данными.и расширил ее.[ . ]
Примером первичной сукцессии является зарастание песчаных дюн или лавовых потоков. Сначала на голых дюнах появляются злаки (растения-пионеры), вслед за злаками — кустарники (ивняк, ольшаник), затем — сосна, а после этого лиственные породы деревьев. Вместе с первыми растениями появляются и первые насекомые (например, кузнечики, как растительноядные) и пауки (как хищники). Потом к первым поселенцам прибавляются муравьи, жуки, бабочки и т.д. Вслед за насекомыми появляются птицы и мелкие млекопитающие, а затем и крупные животные. Развитие, начавшееся в сухом и бесплодном местообитании, заканчивается образованием стабильной лесной экосистемы с мощной, богатой гумусом почвой, с дождевыми червями и моллюсками, разнообразным животным миром. Главную роль в развитии экосистемы играют растения. Вызываемые ими изменения в почве служат основой для смены видового состава сообщества.[ . ]
На рис.6.3-20 показан другой пример интерпретации СЕСЮ1Р в песчаных дюнах, с приведением всех наблюдений и их интерпретации.[ . ]
| Зависимость между числом грызунов и продуктивностью на песчаных равнинах в пустыне Сонора (I и квадраты) и на песчаных дюнах в пустыне Мохаве и Большом Бассейне (II и кружки) [58]. | ![Зависимость между числом грызунов и продуктивностью на песчаных равнинах в пустыне Сонора (I и квадраты) и на песчаных дюнах в пустыне Мохаве и Большом Бассейне (II и кружки) [58].](https://ru-ecology.info/static/pngsmall/820485146.jpg) |
Растения ястребинки Шегаслигп итЬеИаШт в лесу обычно прямостоячие, на песчаных полях — распростертые, а растения с песчаных дюн имеют промежуточный характер. Листья лесных экземпляров самые широкие, экземпляров с дюн — наиболее узкие, а с песчаных полей — промежуточные. Растения с песчаных полей покрыты тонкими волосками — признак, отсутствующий у растений из других местообитаний.[ . ]
| Положение главных пустынь мира относитель- пустынные области; 3 — пустыни с песчаными дюнами; но крупных атмосферных циркуляций и рельефа [948]. 4 — засушливые прибрежные области; 5 — упрощенная 1 — важнейшие горные системы и плато; 2 — крупные схема ориентировки преобладающих ветров. | ![Положение главных пустынь мира относитель- пустынные области; 3 — пустыни с песчаными дюнами; но крупных атмосферных циркуляций и рельефа [948]. 4 — засушливые прибрежные области; 5 — упрощенная 1 — важнейшие горные системы и плато; 2 — крупные схема ориентировки преобладающих ветров.](https://ru-ecology.info/static/pngsmall/882485152.jpg) |
Наконец, ключевым в популяции однолетнего проломника северного (Апйгозасе 8ер1еп1гюпаИз) на песчаных дюнах Польши (динамика представлена на рис. 15.2) оказался фактор Й1—гибель семян в почве, не зависящая от численности растений. В то же время гибель проростков (йг), не будучи ключевым фактором, зависит от плотности (недокомпенсирующий наклон 0,2). У проростков, появившихся из земли первыми, шанс выжить гораздо выше (5утошс1е5, 1977), что предполагает существование напряженной (зависящей от плотности) конкуренции за ресурсы и/или большей уязвимости для различных факторов более мелких, взошедших позднее растений.[ . ]
Эффективность тепловой одышки неодинакова у разных видов и, вероятно, связана с их экологическими особенностями. Выяснено, например, что у серого варана, обитающего на песчаных дюнах, эта реакция более эффективна, чем у шипохвоста, живущего в каменистой пустыне, при меньшей средней и максимальной температурах субстрата.[ . ]
Различают первичную сукцессию — постепенное заселение организмами появившейся девственной суши, оголенной материнской породы (отступившее море или ледник, высохшее озеро, песчаные дюны, голые скалы и застывшая лава после вулканического извержения и т.п.). В этих случаях решающую роль играет процесс почвообразования. Начальное выветривание — разрушение и разрыхление поверхности минеральной основы под действием перепадов температуры и увлажнения — высвобождает или принимает нанос некоторого количества биогенов, которое уже может быть использовано бактериями, лишайниками, а затем и редкой одноярусной пионерной растительностью. Ее появление, а с нею — симбиотрофов и мелких животных значительно ускоряет образование почвы и постепенное заселение территории сериями все более сложных растительных сообществ, все более крупными растениями и животными. Так система постепенно проходит все стадии развития до климаксного состояния.[ . ]
Большинство представителей семейства 2у-gophyllaceae — ксерофитные кустарники или многолетние травы. Они распространены в пустынях тропических и субтропических областей, растут и на песчаных дюнах, пустошах и болотах умеренного пояса.[ . ]
Резко ухудшается экологическая обстановка на морских побережьях. Ураганные волны перемещают массы песка и ила, вследствие чего чистая морская вода становится мутной и грязной. Смываются многие километры песчаных дюн, изменяются очертания береговой линии, в массовом количестве гибнут донные рыбы; крабы, шримсы и песчаные черви уничтожаются почти поголовно, на берег выбрасываются барракуды и акулы.[ . ]
Этот вывод представляется, однако, менее определенным, если принять во внимание пример, заимствованнный размера низкорослого корявого деревца на бедных почвах. В отличие от других елей, она поселяется и на болотах, и на песчаных дюнах, соответственно снижая в этих случаях свои размеры.[ . ]
Считается, что мощный толчок развитию и широкому распространению технологии использования гео синтетиков в виде геотубов — оболочек, заполняемых грунтом методом гидронамыва, дал успешный опыт защиты побережья Атлантик Сити (США, штат Нью-Джерси). В 1993 г. из-за смыва песка в море Атлантик Сити потерял более 30 м территории пляжа. Местами океан «съедал» по 30 см суши в месяц. Проблема встала наиболее остро, когда приливы и штормы полностью смыли пляж и песчаные дюны, что поставило прибрежные здания под угрозу разрушения.[ . ]
Гречишные произрастают в самых разнообразных экологических условиях. В горах они поднимаются до альпийского пояса, например ревень благородный (Шюит поЬПе), который обитает в Восточных Гималаях на высоте 5.100 м над уровнем моря. В холодных арктических пустынях встречается однолетняя кенигия исландская (Костра ЫатНса). Примером приспособления растений к крайне засушливым условиям жизни может служить род джузгун, у видов которого листья редуцированы и их роль выполняют ассимилирующие побеги, частично опадающие после созревания плодов. Некоторые горцы являются водными растениями. Например, в тропических водоемах плавающие острова образованы горцам бородатым (Р. ЬагЪаЬшн), а горец земноводный (Р. атрЫ-Ыит) встречается как в воде, так п на влажных почвах.[ . ]
Хотя каждый знает, что абиотическая среда («физические факторы») контролирует жизнедеятельность организмов, не всегда осознают, что организмы различными способами в свою очередь влияют на абиотическую среду и контролируют ее. Организмы постоянно производят физические и химические изменения инертных веществ, поставляя в среду новые вещества и источники энергии. Химический состав моря и его донных «илов» в значительной степени определяется деятельностью организмов. Растения, обитающие на песчаной дюне, создают почву, совершенно отличную от первоначального субстрата. Коралловые острова южной части Тихого океана — яркий пример того, как организмы влияют на свою абиотическую среду. Из простых исходных веществ, содержащихся в море, в результате деятельности животных (кораллов и др.) и растений построены целые острова. Сам состав нашей атмосферы регулируется организмами, как будет подробно показано в следующем разделе.[ . ]
Всем известно, что абиотическая среда («физические факторы») контролирует деятельность организмов, однако не все понимают, что организмы в свою очередь влияют на абиотическую среду и контролируют ее самыми разнообразными способами. Организмы постоянно изменяют физическую и химическую природу инертных веществ, отдавая в среду новые соединения и источники энергии. Так, состав морской воды и донных илов моря в значительной мере определяется активностью морских организмов. Растения, живущие на песчаной дюне, образуют на ней почву, совершенно отличную от исходного субстрата. Яркий пример того, как организмы изменяют абиотическую среду, — коралловый атолл в южной части Тихого океана. Из простого сырья, предоставляемого морем, животные (кораллы) и растения строят целые острова. Организмы контролируют даже состав нашей атмосферы.[ . ]
Ветровая деятельность. Ветровая эрозия и ветровое переотложение могут иметь важное значение только при незначительном растительном покрове. Поэтому они шире распространены в аридных, семиаридных обстановках или в высокогорных областях. При эрозии из осадка удаляются относительно мелкие частицы, что приводит к дефляции поверхностей террас. Крупные обломки на дефляционной мостовой могут быть обточены и отполированы (вентифакты) и могут быть покрыты пустынным загаром [948]. Материал, выдуваемый ветром, может аккумулироваться здесь же в виде песчаных дюн или переноситься на большие расстояния и отлагаться в виде лёсса [1438,1119,2671] (см. разд. 5.2.8). Ветром отлагается не только мелкозернистый кластический материал, но также может привноситься карбонатный материал, играющий важную роль в почвообразовании.[ . ]
На рис. 15.8 представлена схематическая связь равновесного уровня численности с интенсивностью не зависящего от плотности процесса. Рассмотрим популяцию, в которой рождаемость зависит от плотности, а смертность — только от физических параметров среды, особых в каждой из трех рассматриваемых местностей. На рисунке показаны три равновесные популяции (Nu N2, N3), соответствующие трем уровням смертности, а значит, и трем разным сочетаниям абиотических условий. Например, Уоткинсон и Харпер (Watkinson, Harper, 1978) показали, что различия в численности однолетника Vulpia fasciculata в разных участках песчаных дюн Северного Уэльса связаны в первую очередь с неодинаковой смертностью, не зависящей от плотности. Размножение регулировалось и зависело от плотности, но слабо менялось под действием физических факторов среды (как показано на рис. 15.8).[ . ]
По всей видимости, выедание травы кроликами сдерживало развитие агрессивных, доминантных злаков, позволяя сохраняться относительно высокому флористическому разнообразию. В то же время при усилении интенсивности стравливания видовое богатство лугов может и сокращаться, так как животному приходится переключаться с сильно выеденных предпочтительных видов растений на прочие, еще достаточно обильные. В результате части видов растений может угрожать вымирание. На рис. 19.2 показаны типы зависимости между интенсивностью стравливания кроликами травостоя (по шкале от 0 до 5) и его флористическим богатством для песчаных дюн Фризских островов (Нидерланды).[ . ]
По всей видимости, выедание травы кроликами сдерживало развитие агрессивных, доминантных злаков, позволяя сохраняться относительно высокому флористическому разнообразию. В то же время при усилении интенсивности стравливания видовое богатство лугов может и сокращаться, так как животному приходится переключаться с сильно выеденных предпочтительных видов растений на прочие, еще достаточно обильные. В результате части видов растений может угрожать вымирание. На рис. 19.2 показаны типы зависимости между интенсивностью стравливания кроликами травостоя (по шкале от 0 до 5) и его флористическим богатством для песчаных дюн Фризских островов (Нидерланды).[ . ]
Она представляет собой математическую обработку данных, позволяющую расположить точки на графике таким образом, что те из них, которые соответствуют сообществам, наиболее сходным по составу и соотношению численностей видов, окажутся ближе всего друг к другу. Расстояние между точками, означающими сообщества, сильно различающиеся количественным соотношением видов или видовым составом, будет гораздо больше. Эта работа требует много времени и в настоящее время выполняется исключительно на ЭВМ. Детальное описание методик можно найти в специальных работах (Gauch, 1982). Рис. 16.7, Л демонстрирует ординацию данных (обилие каждого вида) для 50 описаний растительности песчаных дюн Уэльса.[ . ]
Субформации лиственничных лесов характеризуются не только величиной ареала каждого вида лиственницы, но и комплексом экологических и топографических условий, обусловливающих их внутреннюю структуру. Способность лиственницы произрастать на самых различных элементах рельефа и формировать типы леса со своим преобладанием требует объединения этих условий в более крупные однородные единицы — геоморфологические комплексы лиственничных лесов. Геоморфологические комплексы являются таксономической единицей как лесорастительных условий, так и одновременно классификационной единицей типов лиственничных лесов, соответственно равных по рангу и объему. В составе геоморфологического комплекса объединяются типы лиственничных лесов, произрастающие на крупных категориях форм рельефа и характеризующиеся однородностью процессов их формирования. В целом для формации лиственничных лесов Сихотэ-Алиня выделяется 5 геоморфологических комплексов типов леса — высокогорные, горных склонов, горных плато, долин и лиственничных лесов на приморских песках. Последние хотя и занимают ничтожные площади в северной и средней части Сихотэ-Алиня, располагаясь узкой полосой по песчаным дюнам вдоль морского побережья, однако их своеобразные местообитания, связанные с постоянной морской засоленностью цочвы и воздействием морских ветров, и особенности строения позволяют выделить такие насаждения в особый геоморфологический комплекс лиственничных лесов. Весьма характерным является и тот факт, что лиственница в подобных условиях формирует коренные типы леса.[ . ]
Среда обитания растений
Другие очень специализированные растения приспособлены к нарастанию песка, выживая при постоянном закапывании своих побегов за счет очень быстрого вертикального роста. Трава маррам, Ammophila arenaria, специализируется на этом и в значительной степени отвечает за формирование и стабилизацию многих дюн, связывая песчинки вместе. Песчаный пырей Elytrigia juncea также выполняет эту функцию на морском краю дюн и вместе с некоторыми другими первопроходцами, такими как морская ракета Cakile maritima , инициирует процесс строительства дюн, улавливая песок, переносимый ветром.
В ситуациях срастания образуются небольшие насыпи растительности или обломков, смываемых приливом, и они имеют тенденцию увеличиваться по мере того, как скорость ветра падает с подветренной стороны насыпи, позволяя переносящемуся песку (подобранному с береговых берегов) выпадать из воздушного потока. . Растения-первопроходцы физиологически адаптированы к проблемам высокого содержания солей в воздухе и почве и являются хорошими примерами переносчиков стресса, а также обладают некоторыми рудеральными характеристиками.
Экология песчаных дюн - Sand dune ecology
Экология песчаных дюн описывает биологические и физико-химические взаимодействия, характерные для песчаных дюн .
Системы песчаных дюн - отличные места для сохранения биоразнообразия, отчасти потому, что они не очень продуктивны для сельского хозяйства, а отчасти потому, что нарушенные, стрессовые и стабильные среды обитания находятся в непосредственной близости друг от друга. Многие из них находятся под защитой как природные заповедники, а некоторые являются частью более крупных заповедников, включая другие прибрежные среды обитания, такие как солончаки , илистые равнины, луга, кустарники и леса.
Как появляются песчаные дюны

Песчаные дюны выглядят как нечто само собой разумеющееся, но ученые до сих пор не в полной мере понимают, как именно они формируются.
Какие природные механизмы приводят к появлению дюн? Как они остаются стабильными в течение долгого времени? Как они вообще существуют? Эти вопросы являются более сложными, чем кажется на первый взгляд — говорят исследователи.
Изучая снимки из космоса, ученые выяснили, что дюны могут мигрировать. Небольшие дюны могут преодолевать около 100 метров за год, а большие путешествуют несколько медленнее, но, тем не менее, меняют свое местоположение вполне заметно. Если дюны перемещаются, то они должны сталкиваться? Но как это происходит? И почему дюны не сливаются при столкновениях и не приобретают одинаковые размеры?
Как видите, вопросов, связанных с жизнью дюн, довольно много. Ответы, как отмечают ученые, следует искать в самом процессе их формирования. "Наблюдая за образованием дюн, можно выделить два вида песчаных образований. Высота одних составляет около метра, а высота вторых не превышает нескольких сантиметров. Промежуточного размера, что самое интересное, не существует", — рассказывают исследователи, которые построили в пустыне полигон для наблюдения за дюнами, сделав в итоге множество удивительных открытий.
Внутренняя сторона
На внутренней стороне дюн условия менее суровы, и пастбища типа звеньев развиваются с целым рядом пастбищных трав, которые извлекают выгоду из разумного питательного статуса и умеренно высокого pH более стабильных почв, особенно когда в стабилизированных почвах накоплено достаточно гумуса для воды. удержание должно быть улучшено. Такие виды, как овсяница красная и подмаренник, адаптированы для конкуренции друг с другом - за питательные вещества, пространство для выращивания и свет, и известны как растения CSR, т. Е. Имеют в более или менее равных пропорциях характеристики Конкурентов, Стрессоустойчивых и Рудералов.
В старых выбросах могут также быть участки, где грунтовые воды находятся у поверхности и часто поднимаются, вызывая наводнения зимой. Частое, но периодическое переувлажнение корней требует адаптации к стрессу, поэтому здесь увеличиваются доли стрессоустойчивых.
Однако в богатой питательными веществами воде есть некоторые растения с очень конкурентоспособной стратегией, например тростник ( Phragmites australis ). Это пример растения, которое быстро растет и подавляет другие виды, занимая пространство корней и побегов и затеняя оппозицию. Даже его собственные сеянцы не могут прижиться в существующей популяции, но семена разносятся на большие расстояния в больших количествах, чтобы образовались новые колонии, в то время как зрелые популяции расширяются за счет быстрого вегетативного роста боковых подземных побегов - корневищ.
Читайте также: